检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
RNA修饰作为一种关键的表观遗传调控机制已经被广泛研究。迄今为止,已经发现了几百种RNA化学修饰,包括N1-甲基腺嘌呤(m1A)、N6-甲基腺嘌呤(m6A)、5-甲基胞嘧啶(m5C)等。近年来,越来越多的研究揭示:真核生物mRNA的5’末端除了存在典型的m7G加帽修饰外,还存在多种新型的5’端非典型加帽修饰,如烟酰胺腺嘌呤二核苷酸(NAD+)、黄素腺嘌呤二核苷酸(FAD)、 脱磷酸辅酶A(dpCoA)等等。其中NAD+帽子修饰RNA(NAD-capped RNA)已经在不同生物体中被广泛鉴定到,包括在原核生物(如大肠杆菌、枯草杆菌、委内瑞拉链霉菌和沙眼衣原体)、真核生物(如酵母、哺乳动物细胞和植物)以及古细菌中。研究发现,不同生物体中的NAD-RNA种类各异。细菌中的编码蛋白mRNA和一些小型调控RNA、古细菌和真核生物中不同类型的RNA都已经被证明存在NAD+加帽修饰,强调了研究NAD-RNA生物合成、代谢和功能的重要性。
目前已经确定有两类NAD-RNA脱帽酶,分别在原核和真核生物中被相继发现。第一类脱帽酶(Class-I)以大肠杆菌的Nudix水解酶NudC为代表,其可以通过裂解NAD+帽子内部的焦磷酸键,释放烟酰胺单核苷酸(NMN),从而产生一个5’单磷的RNA(p-RNA)。真核生物中的NudC同源蛋白(如酵母中的Npy1和哺乳动物细胞中的Nudt12/16)也已经被证明是NAD-RNA的脱帽酶。第二类脱帽酶(Class-II)是以酵母、拟南芥和哺乳动物中的DXO/Rai1家族酶为代表的一类酶,其可去除整个NAD+帽修饰,生成p-RNA。酵母中的5’-3’外切核酸酶Xrn1和Rat1也可以以同样的方式去除NAD+帽子修饰,但主要作用于线粒体NAD-RNA。然而,NudC和DXO/Rai1两类酶也可作用于FAD帽子修饰和dpCoA帽子修饰的RNA,并非特异地作用于NAD-RNA。

2024年3月15日,北京大学生命科学学院和北大-清华生命科学联合中心陈雪梅课题组在Nature Communications发表了题为Toll/interleukin-1 receptor(TIR) domain-containing proteins have NAD-RNA decapping activity的研究论文,该研究首次发现来自细菌和古细菌的TIR结构域蛋白具有强烈的NAD-RNA脱帽酶活性,能够从NAD+帽子修饰中去除NAM基团,产生一种新的非典型加帽修饰RNA(即环化ADPR-RNA或者其变体)。同时,研究还发现该TIR结构域蛋白介导的NAD-RNA脱帽活性高度依赖于谷氨酸残基催化位点,且在促进TIR结构域蛋白寡聚的条件下表现出增强的活性。TIR结构域蛋白显示出特异于其他已知脱帽酶的性质,即可以特异性地作用于NAD-RNA,而不作用于其他非典型帽子修饰的RNA。
研究进一步证明,AbTir在细菌体内具有功能——在大肠杆菌中诱导表达AbTir蛋白可抑制细胞生长,显著降低游离NAD+和NAD-RNA的水平。全转录组范围内的NAD-RNA测序分析表明,大肠杆菌中一小部分涉及“分子运输”和“氧化还原酶活性”的基因产生的NAD-RNA易受到AbTir的靶向。本研究首次证明了TIR结构域蛋白具有NAD-RNA脱帽活性,提示它们在体内可能也参与NAD-RNA的动态调控,为后续研究NAD-RNA的生物学功能提供了良好的基础。有趣的是,TIR结构域蛋白被频繁报道参与生物体的抗病免疫过程,然而其背后的作用机制尚不完全清楚。本研究结果暗示TIR结构域蛋白可能通过影响NAD-RNA的表达水平在生物学抗病免疫中发挥作用,进一步的研究将有助于挖掘生物体抗病免疫新机制。
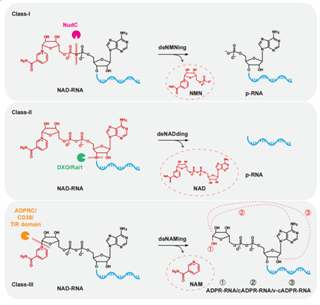
图1: 三种不同的NAD-RNA脱帽机制总结。
北京大学生命科学学院助理研究员王旭峰博士、德国科隆大学生物化学研究所于东立博士(现为美国波士顿哈佛医学院Dana-Farber癌症研究所博士后)为论文共同第一作者;北京大学生命科学学院陈雪梅教授为论文通讯作者;该工作还得到了美国犹他大学医学院Qi Chen教授和西湖大学柴继杰教授的帮助和指导。该研究得到了National Institutes of Health (NIH)、科技部国家重点研发计划项目的资助。
地址:北京市海淀区颐和园路5号
金光生命科学大楼
电话:010-62757794

北大生科官方微信

生声不息公众号