检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
2023年夏天全球平均气温再创新记录,高温频次也逐年增加。刚过去的北京夏天的高温天气令人记忆犹新,最高气温超过35℃的高温日数近一个月,是常年高温天数的3倍多。高温强度和频次的增加不仅造成严重的粮食减产和世界粮食安全问题,也使动植物的生存面临严峻挑战。高温已成为科学家和各国政要们共同关注的人类可持续发展问题。与动物不同,植物不能通过移动避开高温,但植物可通过可塑性发育即热形态建成来提高对高温的适应性。动物虽然能通过移动避开高温,但动物的卵却不能移动,其孵化温度高低常会决定动物的性别,如在较低温度下(26℃)孵化出的巴西龟全部为雄性,而在较高温度下(32℃)则全部为雌性。虽然在过去十几年中,科学家对植物热形态建成的分子机制已有较深入的研究,但高温如何影响植物器官的命运决定却少有报道。
2023年9月13日,北京大学生命科学学院秦跟基教授课题组在Nature子刊Nature Communications上在线发表了题为“Arabidopsis TCP4 transcription factor inhibits high temperature induced homeotic conversion of ovules”的研究论文,揭示了重要转录因子TCP功能冗余地在高温下保护植物胚珠命运不向心皮同源转化的分子机制。
植物的胚珠是种子的前身,不仅是植物进行有性生殖传播下一代的基础,也是农业生产中主要收获器官,对粮食安全至关重要。作为花中第四轮器官,胚珠的器官形成和决定机制一直是科学家感兴趣的科学问题。根据ABCDE花发育模型,第三轮器官心皮(组成雌蕊)的命运由MADS-box转录因子AG-SEP形成的复合体来控制,而被心皮包裹的第四轮器官胚珠则由MADS-box转录因子AG-SEP-STK/SHP复合体来决定。这样出现一个一直以来被忽视的科学问题,也就是既然四类MADS-box转录因子AG、SEP、STK和SHP均存在于胚珠中,AG-SEP复合体和AG-SEP-STK/SHP复合体在胚珠中就共存,那在胚珠中AG-SEP复合体的功能如何被抑制,从而使AG-SEP-STK/SHP复合体功能起主导作用来促进胚珠命运正常形成的呢?
秦跟基教授课题组长期研究植物重要转录因子家族TCP的功能,经过近十年的努力,通过遗传杂交和CRISPR/Cas9定点敲除技术,该研究团队构建了一个敲除Class II TCP亚家族11个成员及TCP16功能的十二重tcp突变体tcpDUO(tcp duodecuple mutant, tcp2/3/4/5/10/13/17/24/1/12/18/16)。十二重突变体tcpDUO具有严重的育性缺陷表型,正反交遗传实验和表型观察证明是雌配子体的发育异常导致该突变体育性缺陷,而花粉完全正常。很有意思的是,在22℃下,突变体tcpDUO的胚珠命运以较低频率(0.2%)向心皮样结构同源转化(图1a,b),而在28℃高温条件下,突变体tcpDUO的胚珠全部转变为心皮样结构(图1c,d,f,g),在tcpDUO中表达TCP4可互补其胚珠同源转化的表型(图1e)。说明TCP在高温条件下具有保护胚珠命运不向心皮转变的重要功能。
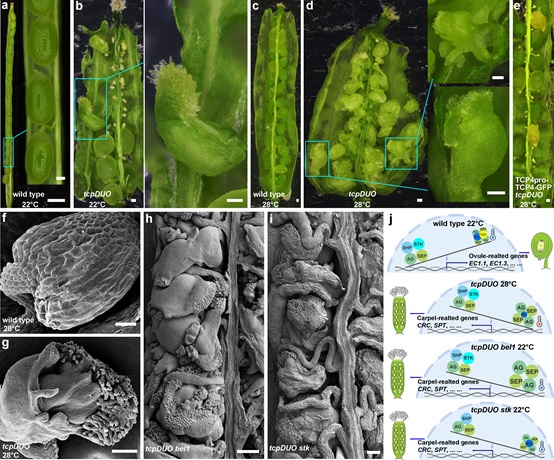
图1. TCP转录因子在高温下保护胚珠命运不被转变。
a. 在22℃生长条件下野生型的胚珠。b. 在22℃条件下十二重突变体tcpDUO的胚珠有极个别转变为心皮样结构。c. 在28℃条件下野生型的胚珠没有转变为心皮的现象。d. 在28℃条件下十二重突变体tcpDUO的胚珠全部同源转化为心皮样结构。e. 表达TCP4可互补tcpDUO的胚珠在28℃条件下全部同源转化为心皮样结构的表型。 f. 在28℃条件下野生型的胚珠扫描电镜图。g. 在28℃条件下十二重突变体tcpDUO的胚珠转变成的心皮样结构扫描电镜图,清楚可见心皮特有的乳突状细胞。h. 十三重突变体tcpDUO bel1在22℃条件下全部转变为心皮样结构。i. 十三重突变体tcpDUO stk在22℃条件下也全部转变为心皮样结构。j. TCP转录因子控制胚珠命运的工作模型。野生型22℃条件下,TCP和BEL1共同抑制AG-SEP复合体功能,使得AG-SEP-STK/SHP复合体起主导作用,从而决定了胚珠命运;野生型在28℃条件下,BEL1蛋白虽然部分降解,但TCP已可充分抑制AG-SEP3的功能,从而保护胚珠命运不被转变;tcpDUO 在28℃条件下,TCP功能缺乏,同时BEL1蛋白部分降解,无法充分抑制AG-SEP的功能,使胚珠全部转变为心皮样结构;在tcpDUO bel1中,即使在常温下,由于同时缺乏TCP和BEL1的功能,使得AG-SEP复合体功能不被抑制,从而起主导作用,使胚珠全部转变为心皮样结构;而在tcpDUO stk中,AG-SEP-STK/SHP复合体功能部分缺失,同时TCP功能缺乏,无法充分抑制AG-SEP复合体功能,使得AG-SEP复合体功能起主导作用,导致胚珠全部转变为心皮样结构。
为解析TCP保护胚珠命运的分子机制,通过去雄后收集未授粉的雌蕊进行转录组测序和胚珠的RT-qPCR分析,确定了tcpDUO的胚珠中控制心皮发育的关键基因同时也是AG-SEP复合体直接下游因子CRC和SPT在胚珠中异位表达。通过多种生化实验确定了TCP4蛋白与AG以及SEP3相互作用,抑制AG-SEP3的转录激活活性,从而抑制其下游CRC和SPT等在胚珠中的异位表达,促进胚珠命运正常形成。BEL1是抑制AG-SEP复合体功能的已知转录因子,该研究进一步确定了BEL1蛋白在高温下通过26S蛋白酶体降解,而TCP4通过与BEL1蛋白互作抑制其降解,从而促进了BEL1对AG-SEP复合体功能的抑制。通过在tcpDUO背景下敲除BEL1或STK构建tcpDUO bel1或tcpDUO stk十三重突变体,发现tcpDUO bel1或tcpDUO stk的胚珠在正常生长温度下全部转变为心皮样结构(图1h,i),说明TCP在胚珠命运决定过程中与BEL1及STK具有功能冗余性,是胚珠命运决定的关键正调控因子。
该研究不仅通过扎实的遗传学数据证明了TCP转录因子在胚珠命运决定中的新功能,还阐明了植物胚珠命运在高温下的不稳定性以及TCP在高温下通过跷跷板模型保护胚珠命运不被改变的分子机制。在该跷跷板模型中,TCP转录因子通过与决定心皮命运的AG-SEP3复合体直接相互作用并抑制其功能,同时还通过与转录因子BEL1直接相互作用抑制其在高温下的降解,从而加强了BEL1对AG-SEP3复合体功能的抑制,进而保证了AG-SEP-STK/SHP复合体在胚珠中起主导作用来决定胚珠的正常命运(图1j)。
北京大学生命科学学院已毕业的兰婧秋博士(现为中国科学院遗传与发育生物学研究所曹晓风院士课题组博士后)和博士研究生王宁为该论文的共同第一作者,北京大学生命科学学院秦跟基教授为通讯作者,北京大学生命科学学院博士后王宇涛、博士研究生姜一丹及已毕业的于浩为共同作者,中国科学院遗传与发育生物学研究所曹晓风院士在研究中给予了重要支持和指导。该研究得到国家自然科学基金(31970194)、国家杰出青年科学基金(31725005)、国家自然科学基金创新群体(31621001)以及北京大学蛋白质与植物基因研究国家重点实验室的资助。
地址:北京市海淀区颐和园路5号
金光生命科学大楼
电话:010-62757794

北大生科官方微信

生声不息公众号